Research Article
Suliman A.I.Ali1*, Mory Mandiana Diakite1, Saqib Ali1, and Man-Qun Wang1
1Hubei Insect Resources Utilization and Sustainable Pest Management Key Laboratory, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, People?s Republic of China
Corresponding author
Suliman A.I. Ali, Hubei Insect Resources Utilization and Sustainable Pest Management Key Laboratory, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, People?s Republic of China, E-mail: suliman@webmail.hzau.edu.cn
Received Date: 02nd April 2015
Accepted Date: 01st September 2015
Published Date: 05th September 2015
Citation
Ali SAI, Diakite MM, Ali S, Wang MQ (2015) Understanding Insect Behaviors and Olfactory Signal Transduction. Enliven: J Genet Mol Cell Biol 2(2): 004.
Copyright
@ 2015 Dr. Man-Qun Wang. This is an Open Access article published and distributed under the terms of the Creative Commons Attribution License, that permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Olfaction is a molecular sense, in which information carried in airborne chemicals is transformed into patterns of brain activity that underlie odor
perception. It is probably the most important sense for survival of most animal species ranging from insects to mammals. Detection and localization of
food, avoidance of toxins and predators, and communication with cohorts and mating partners through volatile pheromones are examples of the range
of olfaction dependent behavior. Olfaction in insect, both medical and agricultural fields, is well documented today. That wide knowledge of insect
olfaction (behaviour), especially in agriculture, has contributed to the development of the Integrated Pest Management (IPM) strategies, especially the
use of semiochemicals for luring, trapping and killing of insect pests. The present literature review addresses the following general subjects: notion of
the pest, importance of the sensilla in insect life, general mechanism of chemical signals transduction, odor identification and discrimination in insects,
chemical messengers (pheromones and allelochemicals), insect-plant interactions, ratio-specific odor recognition, and evolution in pest-control strategies
and the use of plant extracts in protecting stored-products. The purpose is to contribute in helping beginners in the modern agricultural entomology field
for rapid familiarization with these terms frequently used in IPM strategies.
Keywords:
Sensilla; Olfactory organs; Signal transduction; Predator and parasite
Introduction
Insect relies on chemosensory or chemoreceptor organs located on antennae, mouthparts, wings, legs and ovipositors to live. In general, the chemosensory includes gustatory receptors involved in sense of taste and olfactory receptors involved in sense of odor. In other words, the olfactory receptors are most abundant on the antennae, but may also be associated with the mouthparts or external genitalia (especially near the tip of the female's ovipositor); while the taste receptors are most abundant on the mouthparts, but may also be found on the antennae, tarsi, and genitalia. The gustatory receptors are commonly described as thick-walled hairs, pegs, or pits; whereas olfactory receptors are usually thin-walled pegs, cones, or plates with numerous pores. Anyway, in either case the odor molecules enter through the openings (pores) located on the cuticle where the dendrites of several (usually up to five) sensory neurons are exposed. In sensory neuron cells, there are receptor genes encoding proteins, which mediate odor signal transduction. These small soluble proteins called OBPs (Odorant Binding Proteins) are secreted in large quantities by support cells surrounding the Olfactory Sensory Neurons (OSNs) [1]. They bind odorant messages allowing therefore, an insect to locate food source, aggregate, and mate [2-4].
Notion of the Pest
In agriculture, an insect species is considered as Pest if it can cause important damage to the growing crops or stored-products or livestock production. Thus, because of the competition with human beings for staple food, hundreds of insect species are classified as pests. They feed on leaves or burrows in stems, fruits, roots and stored grains. Those who depend on growing crops (e.g. green leaves) to live are called phytophagous; whereas those who feed on grains, especially the stored-grains, are called stored-grain pests or post-harvest pests. In terms of preference for food (host), the phytophagous, in turn, can be categorized into three groups, of which: monophagous (feeding on plants within a single genus), oligophagous (having hosts in different genera within the same plant family) and polyphagous (attacking a large number of plants of different families) [5,6]. At a very specific level, the insects can be categorized as leaf feeders, stem feeders or grain feeders, etc. Additionally, some insect pests are aggressive only when instar (case of lepidopteran pests), while others are dangerous in both larval and imago stages. The second type of damages, which makes an insect species as pest, is the transmission of epidemic diseases (e.g., bacterial, viral, or fungal infection) to the crops or facilitation of plant infection by epidemic diseases. However, beyond the damages they cause on crops and stored-products, the insects also play many important roles in the nature. For instance, they aid bacteria, fungi, and other organisms in the decomposition of organic matter, in soil formation as well as plant pollination [7-10]. Certain insects provide sources of commercially important products such as honey, silk, wax, dyes, or pigments, all of which can be of direct benefit to man. Other species like grasshopper plagues, termite swarms, large palm weevil grubs, etc are still sources of protein in some countries. In the biological control of the insect pest, many insect species are used as predators or parasites. Briefly, the word ?Pest? is a variable notion. A species becomes a pest if it appears where it?s unwished or competes with human beings for staple foods.
Sensilla in Insect Survivorship
The sensilla are usually small hairs within which are housed olfactory organs adapted for perception of specific stimuli (e.g., touch, smell, taste, heat, cold) [11-15]. In most insects, Odorant receptors (ORs), gustatory receptors (GRs) and ionotrophic receptors (IRs) are olfactory organs (Figure 1c) mediating the binding of external odor molecules. Although these small sense organs occur all over the body surface, they are particularly abundant in antennae, palps, and cerci (cerci are sometimes called pincers). Antennae are the main organs bearing olfactory receptors [6,16,17]. They play the dominant role in insect?s olfaction because they are more exposed to air currents as insects move upwind toward an odor source [18-20]. Depending on the external morphology, the antennae can be classified into different types and subtypes. Sensilla trichodea, sensilla placodea, sensilla basiconica, sensilla chaetica, sensilla coeleoconica, sensilla styloconica and sensilla campaniformia are among of the well-characterized sense receptor types [21]. The sensilla can be hairs, pegs, plugged or open grooves. They can be on the surface, or they can be located within a depression or a pit with a restricted opening [22,23]. The same sensilla type can be found in different species with the same characteristics. But in some cases, it can have some difference between sensilla in different species even being the same type. For example, a sinsillum can have smooth surface in a given species, while in other species, it can be grooved or striate or pitted [24-26]. The sensillum types present on moth antennae can be classified into six different groups. These are the sensilla trichodea, sensilla basiconica, sensilla auricillica, sensilla chaetica, sensilla styloconica and sensilla coeloconica. Sensilla trichodea are very long, thin, porous hair-like structures with sharp pointed tips. Sensilla can be categorized according to their putative function as well, of which some are mechanoreceptors (campaniformia and sensilla chaetica) or chemoreceptors (sensilla basiconica) or hygro or thermosensitive. A same type can even be mechano and chemoreceptors at the same time (sensilla digitiformia). Sensilla digitiformia is also thought to be hygroreceptor because of their abundance on the mouthparts of some hygrophilous species that depend on damp or wet patchy biotopes/seasons [27]. S. trichodea are known to play a major role in sensing mechanical stimuli and in detecting chemical stimuli in insects [25,26,28,29], and are thought to be innervated by 1?3 sensory cells [30], while sensilla coeloconica are thermosensitive and innervated by 1?5 sensory cells [31-33]. S. basiconica are considered to be olfactory receptors in phytophagous insects [25,34]. The morphology of this sensilla suggests that they are sensitive to many kinds of chemical stimulus [20,35-37]. Additionally, S. cavity is thought to be involved in perception of humidity and temperature [23] and might play a role in preventing desiccation [38]. Both olfaction and contact chemoreception are involved in host-plant selection, as well as in the integrity of insect societies, especially in nestmate recognition [25]. In sensilla characterization, it can have sexual dimorphism [20,39]. The ability of recognizing and discriminating thousands of odorant molecules in insects as in mammals (Figure 2) relies on specialized chemosensitive neural cells to express olfactory receptor proteins, which reside within segregated compartments called sensilla [40-42]. Lots of sensilla are located on the surface of the insect antennae with olfactory neurons being protected inside the binding of a ligand to a receptor protein is the key event in olfactory transduction, as it converts a chemical signal in the environment into an electrical signal that can be interpreted by the insect nervous system.
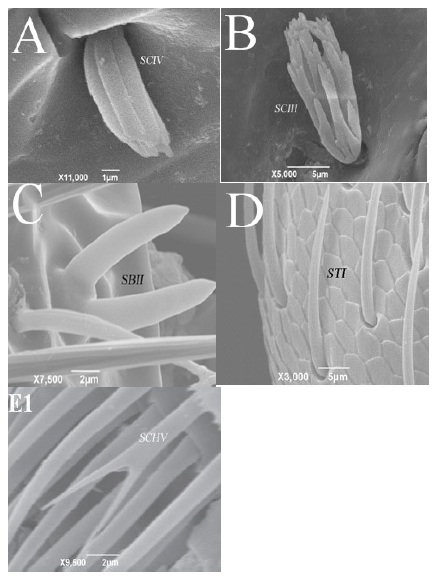
Figure 1: sensilla types A and B are sensilla from Sitophilus granarious which are classified as Sensilla coeloconica, while C, D and E are of Tribolium castaneum, and classified as Sensisilla basiconica, S. trichodea and Sensilla chaetica.

Figure 2 : Odorant Receptors and the Organization of the Olfactory System [125].
Mechanism of Chemical Signals Transduction
The mechanism starts by trapping chemical molecules dispersed in the air by special chemoreceptors or olfactory receptors expressed in the cell membranes of olfactory receptor neurons. Activated olfactory receptors initiate a signal transduction cascade which ultimately produces a nerve impulse (or action potential or spike). The neurons (electrically excitable cells) then processed and use neurotransmitter (an endogenous chemical stored in synaptic vesicle) to transmit signals from one neuron to the next, usually from the axon terminal till the signals reach central nervous system. In other words, axon terminals (distal terminations of the branches of an axon) are separated from neighboring neurons by a small gap called a synapse [43,44], across which impulses or signals are sent before being received by postsynaptic receptor and in a pattern neuron-to-neuron via synapses, the external signals converted into internal electrical signals or impulses by sensory neurons (Figure 3b), ultimately reach the spinal cord and brain (the two constitutive components of the central nervous system). Multipolar neurons play an important role in this complex neural communication or cell-to-cell communication. They allow for the integration of a great deal of information from other neurons. They include motor neurons and interneurons, and constitute the majority of neurons in the brain. We have to remember that second messengers intervene in this process by relaying signals, causing some kind of change in the activity of the cell and amplifying the strength of the signals that will be sent to the brain through glomeruli. They are a component of signal transduction cascades. In humans as well as most vertebrates, the sense of smell is mediated by specialized sensory cells of the nasal cavity (Figure 2), which can be considered analogous to sensory cells of the antennae of invertebrates (Figure 3) [45].

Figure 3: Anatomy of an olfactory sensillum [139] and Summary of odorant sensitivities and odorant receptor expression in different types of sensilla (Maria and Liqun, 2008, [13]).
Odor Identification and Discrimination in Insects
In insects, each sensillum (Figure 3a) possesses specialized sensory cells called ORNs that run chemical signals inward to the central nervous system. The individual ORNs express specific type of odorant receptor proteins called OBPs whose role is to bind external odorant (Figure 3b) and initiate transduction. After binding the odorant, the activated receptors send transduction signal (Figure 3b) to the glomeruli [46,47] via second messengers. We have to precise that each glomerulus receives signals from multiple receptors that detect similar odorant features. Because multiple receptor types are activated due to the different chemical features of the odorant, multiple glomeruli will be activated as well. All of the signals from the glomeruli will then be sent to the brain, where the combination of glomeruli activation will encode the different chemical features of the odorant. The brain will then essentially put the pieces of the activation pattern back together in order to identify and perceive the odorant [46,48,49]. Odorants that are alike in structure activate related patterns of glomeruli, which lead to a similar perception in the brain [46,50-53]. Within the glomerular array, the synaptic organization of afferent ORN axons and dendrites of antennal lobe interneurons forms the mechanism underlying odor identification and discrimination [54-56]. Generally, each OSN expresses one (or sometimes a few) OR and OSNs that express the same OR converge on a single glomerulus in each hemisphere [48,57-61]. Anyway, studies showed that OSNs may innervate the entire glomerulus, or just the glomerular periphery. In bees, OSNs from the distal antennal segments innervate the outer layer of the glomerular cap, and more proximal OSNs innervate the central layers [62]. The current dogma is that axons from all ORNs expressing the same receptor converge onto one or two glomeruli of a possible 1800 glomeruli in each olfactory bulb [63]. We have to remember that the glomeruli are located near the surface of the olfactory bulb (forebrain involved in odor perception). And, a glomerulus is the basic unit in the odor map of the olfactory bulb. The primary olfactory brain center of insects, the Antennal Lobe (AL), constitutes the first synaptic relay station of the antennal afferent pathways, as it receives input from antennal olfactory sensory neurons and sends the output to higher brain centers [64-67]. The building blocks of the AL found in most insect orders are the olfactory glomeruli, in which the interactions between antennal and deutocerebral neurons take place [68]. Additionally, the real difference between an insect's sense of smell and sense of taste lies in the form of the chemical it is collecting. Smelling is going with odor, while tasting occurs if the chemical substance is in solid or liquid status.
Chemical Messengers
Pheromones
Pheromones are chemical signals that are secreted by an animal to the outside and cause a specific reaction in a receiving individual of the same species [69]. They consist in long carbon chains derived from the metabolism of fatty acids [70]. The pheromones usually wind borne, but may be placed on soil, vegetation or various items [71,72] as messengers. As reported earlier, the transport of the volatiles or pheromones from the external environment to the olfactory receptors is mediated by small proteins, called OBPs [73-75] with 135?220 amino acids long [75,76]. The bound odor molecules are then processed and conveyed to the central nervous system which produces all behavioral responses thereafter on receiving species [6,16,17]. The olfactory receptor genes encoding OBPs can be divided into Pheromone-Binding Proteins (PBPs) involved in the recognition of sex pheromones and General Odorant Binding Proteins (GOBPs) which are thought to participate in the recognition of general odorants [13]. Like OBPs, coding sequence of PBPs shows the six conserved cysteine residue's position linked by three disulfide bonds [77]. Chemosensory Proteins (CSP) constitute another class of small binding proteins. They are more conserved comparatively to OBPs, and are characterized by the presence of 4 conserved cysteines that form two disulfide bridges [78]. They may evolve from the OBPs in the early development of arthropods [79].
Types of Pheromones
There are different types of pheromones according to the response they induce in the Perceiving Individuals [80]. The most common pheromones are: Sex pheromones (usually emitted by females to induce mating behavior in males), Aggregation pheromones (usually secreted by males to induce host finding behavior, defense against predators or overcome host resistance by mass attack), Alarm pheromones (triggered once there is a threat), Trail pheromones (specific in social colonies to indicate the way leading to the discovered-food source), and lastly, Host marking pheromones (produced to reduce the competition between members of the same species) [70;81]. We have to precise that male-produced sex attractants have been called aggregation pheromones, because they usually result in the arrival of both sexes at a calling site; whereas most sex pheromones are produced by the females.
Allelochemicals
In chemical ecology field, pheromones and allelochemicals are both used, whence the reason to make the difference between the two terms. In other words, like pheromones, the allelochemicals are biochemical produced to influence the growth, survival, and reproduction of other organisms (Allelopathy). They can have beneficial (positive allelopathy) or detrimental effects (negative allelopathy) on the emitter or the attacker [82]. As shown in (Figure 4), the allelochemicals can be classified into two groups considering whether they act as intraspecific (pheromones) or interspecific (allelochemicals) mediators. Allelochemicals include allomones (emitter species benefits), kairomones (receptor species benefits) and synomones (both species benefit) [83]. Basically, the volatiles released by plants (Figure 5) can be categorized into four groups as followed [6]:
? Attractant: a chemical that causes an insect to make oriented movements toward the source of the stimulus.
? Repellent: a chemical that causes an insect to make oriented movements away from the source.
? Feeding or oviposition stimulant: a chemical that induces feeding or oviposition behavior.
? Deterrent or antifeeding stimulant: a chemical that inhibits feeding or oviposition.
However, a single chemical signal may act as both as pheromone and allelochemical, as attractant as repellent.

Figure 4:Types of semiochemicals

Figure 5:Biosynthetic pathways leading to the release of plant volatiles.
Insect-Plant Interactions
Interactions between plants and insects can be both antagonistic and mutualistic [84,85]. The plant volatiles are signals used by insects to locate hosts, find mates, prey and select oviposition sites [86]. The majority of volatiles collected from plants, and grains are: terpenoids, fatty acid derivatives, benzenoids and nitrogen-containing compounds. Some of the volatiles are specific to certain species, while others are general and found in many species [87,88]. Once released by a source (Figure 5), the volatiles are dispersed (Figure 6), mixed, and diluted by the ambient motion of air to form a shifting and ?lamentous plume [89]. The relationship between insects and plants is largely influenced by the ambient air that disperses odor molecules. The olfactory world is characterized by constant movement and ?ux. Gradients of plant odors do not concentrate a few centimeters distant from a plant due to air turbulence [6]. Explicitly, host odors exist in the form of pockets blowing downwind along with the non-odorous air creating an odor gradient. As the wind swings about, the order is broken into a series of pockets (Figure 6). And, by positive anemotaxis the insects will identify and reach a food source [90]. Indole, a product of the shikimic acid pathway, is formed from indole-3-glycerol-P either as an intermediate in Trp biosynthesis or by a Trp-independent pathway leading to a family of nitrogen-containing defense compounds (e.g. 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one) [91]. Sesquiterpenes are synthesized via the isopentenyl pyrophosphate (IPP) intermediate following the classical mevalonate pathway, whereas monoterpenes and diterpenes are synthesized via an alternative IPP pathway with glyceraldehyde-3-P and pyruvate identified as the direct precursors of IPP [92]. The mevalonate pathway is localized in the cytosol and reactions to the non-mevalonate pathway are localized in plastids. The homoterpene (E) -4,8-dimethyl-1,3,7-nonatriene and (E, E) -4,8,12-trimethyl-1,3,7,11-tridecatetraene are derived from their 15 and 20 carbon precursors, farnesyl- and geranylgeranyl-pyrophosphate, respectively, by a series of enzymatic steps with the overall loss of four carbon units [93]. The green-leaf volatiles derive from linolenic acid via a 13-hydroperoxylinolenic acid intermediate [94]. This oxidized linolenic acid, instead of losing water and committing the molecule down the defense signaling jasmonic acid pathway, is cleaved to form two fragments of 12 and six carbon units (Figure 3). The variety of green-leaf volatiles are formed from this second pathway by multiple rearrangement steps of the six-carbon (Z) -3-hexenal [95].

Figure6: Dispersal of odour source [6].
Ratio-Specific Odor Recognition
Numerous electrophysiological studies on a wide range of phytophagous insects have demonstrated that peripheral receptors are tuned to the detection of ubiquitous plant volatiles. Because phytophagous insects generally recognize the host odor by using ratios of common plant volatiles, and recognition is thus not restricted to species-specific compounds, it would appear that the central processing of peripheral signals is extremely important [88]. Many insect studies have suggested that generalist olfactory receptors are responsible for host plant odor perception, but there is some evidence for the presence of specialist olfactory receptors for the detection of plant odor in insects such as Spodoptera littoralis. Pheromone detectors usually are very sensitive and selective to their specific key compounds. The neurons are likely to function as labeled lines, with each type of neuron carrying information about a single odorant compound [96]. Chemical cues released into the air can guide moths (nocturnal flyers) to food sources or mating sites over long distances [97], even at very low concentrations (a few molecules of pheromone or plant odors). Different flower species often share many volatile components [88,98], but their combination and concentration are unique to each species, forming an ?odour code? [99] that specialist insect may use to identify their hosts. Ratorderf compounds are assumed to drive host plant location in insects [16,88,100]. Even the lowest level of a compound in a blend might contribute to the attraction of an insect species to its host plant [101,102].
Evolution in Pest-Control Strategies
The primary objective for pest control was to avoid the spread of diseases by insects through land managing, improved housing or sanitation of food facilitation before storing. Indeed, the continuous effort for controlling pest led early to the production of DDT, a remarkable compound that is highly toxic to most insects and long-lasting in effect. Widely used in agriculture for many years, DDT is not anymore an ideal insecticide because of environmental concerns. Similar ecological problems were encountered with many successors to DDT like Dieldrin or Endrin. In other words, the continuous use of synthetic insecticides, in particular, has not only caused death through poisoning, accumulated in man, concentrated in food chains, but also caused resistance in pest populations and destroyed parasites, predators and pollinators. Thus, biological methods as well as the use of semiochemicals becomes increasingly important with the increase of the restriction on the use of undesirable insecticides [103]. We have to remember that, the biological methods include introducing pest strains that carry lethal genes or parasitoids, while semiochemicals consist of the use of pheromones to trap and disrupt mating behavior of the insect pests. The role of feeding-induced plant volatiles in host habitat location by natural enemies is well documented [85,104-107]. Exploitation of plant volatiles that attract natural enemies has been potential for enhancing biological control in agroecosystems [108]. For instance, synthetic herbivore-induced plant volatiles (HIPVs) have been used to attract and retain beneficial insects into vineyards and hopyards [109,110]. However, a successfully pest management requires combined strategies for an adequate long-term solution [111]. And, the presence of some insect pests in the field does not automatically result in damage or yield loss. Therefore, there is a threshold that should be reached before deciding treatment.
Pheromones and Semiochemicals in Pest Control
The characterization of the first insect sex pheromone was established by [112], and was isolated from female Bombyx mori (Lepidoptera). This discorvery led to the development of commercial activities in the synthesis of semiochemicals previously identified as potential agents for controlling pests. Since then we have assisted the replacement of synthetic insecticides with pheromone products [81,97], and the emergence of a new scientific discipline: the chemical ecology or Chemical communication. In parallel, the gas chromatography appeared in chemistry and brought simplicity in identification of volatile molecules. Rapidly, the economic interest for using pheromone compounds in pest controls was updated and included in Integrated Pest Management (IPM) programs [70], which imply various strategies depending the goals and scopes to achieve. Monitoring of insect populations, trapping by using traps lured with synthetic attractant associated with a killing substance [113], Push-pull strategy, which consists of pushing away crop enemies while luring them to the pheromone dispensers containing killing substances, is among of the current IPM strategies [114-118]. We have to precise that the aggregation pheromones are one of the most ecologically selective pest suppression methods. They are nontoxic and effective at very low concentrations [119]. An Additionally, crop rotations, field sanitation (crop residue management), seed quality, weed management, tillage, healthy soil, crop and variety selection, intercropping etc, are also part of integrated pest management strategies, especially in organic crop systems.
Conclusion
Understanding the technical terms relevant to insect behavior and chemical messenger?s transduction could be of paramount importance for beginners in the entomology field. All these terms addressed to here have been already previously reported in different sources. However, the effort provided here to collect and define and differentiate the similar terms in an easy way could be very useful for many people in the modern agricultural entomology field in which the terms such as semiochemicals, are frequently used.
References
- Swarup S, Williams TI, Anholt RR (2011) Functional dissection of Odorant binding protein genes in Drosophila melanogaster. Genes Brain Behav 10: 648-657.
- Wyatt TD (2003) Pheromones and Animal Behaviour: Communication by Smell and Taste. Oxford Univ Press.
- Sun YL, Huang LQ, Pelosi P, Wang CZ (2012) Expression in Antennae and Reproductive Organs Suggests a Dual Role of an Odorant-Binding Protein in Two Sibling Helicoverpa Species. PLoS One 7: e30040.
- Pelosi P (1998) Odorant-binding proteins: structural aspects. Annals of the New York Academy of Sciences 855: 281-293.
- Hsiao TH (1985) Feeding behaviour. In Comprehensive insect physiology. biochemistry and pharmacology: Kerkut, G.A.and Gilbert, LI (Eds), Pergamon Press Ltd 9: 471-512.
- Bernays EA, Chapman RE (1994) Host-plant Selection by Phytophagous Insects. Chapman and Hall, New York 312.
- Hill PSM, Wells PH, Wells H (1997) Spontaneous flower constancy and learning in honey bees as a function of colour. Anim Behav 54: 615-627.
- Stout JC, Allen JA, Goulson D (1998) The influence of relative plant density and floral morphological complexity on the behaviour of bumblebees. Oecologia 117: 543-550.
- Chittka L, Gumbert A, Kunze J (1997) Foraging dynamics of bumble bees: correlates of movement within and between plant species. Behav Ecol 8: 239-249.
- Goulson D, Ollerton J, Sluman C (1997) Foraging strategies in the small skipper butterfly, Thymelicus flavus: when to switch? Animal Behav 53: 1009-1016.
- Schlyter F, Birgersson GA (1999) Forest beetles. In Pheromones of non Lepidopteran insects associated with agricultural plants. Oxford: CAB International 113-148.
- de Bruyne M, Baker TC (2008) Odor detection in insects: volatile codes. J Chem Ecol 34: 882-897.
- Benton R, Vannice KS, Gomez-Diaz C, Vosshall LB (2009) Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 136: 149-162.
- Ghaninia M, Ignell R, Hansson BS (2007) Functional classification and central nervous projections of olfactory receptor neurons housed in antennal trichoid sensilla of female yellow fever mosquitoes, Aedes aegypti. Eur J Neurosci 26: 1611-1623.
- Bruce TJ, Pickett JA (2011) Perception of plant volatile blends by herbivorous insects ? Finding the right mix. Phytochemistry 72: 1605-1611.
- Visser JH (1986) Host odour perception in phytophagous insects. Ann Rev Entomol 31: 121-144.
- Panda N, Khush GS (1995) Host plant resistance to insects. CAB International 431.
- Gewecke M, Heinzel HG, Phillippen J (1974) Role of antennae of the dragonfly Orthetrum cancellatum in flight control. Nature 249: 584-585.
- Chapman RF (1998) The Insects Structure and Function. fourth ed. Cambridge University Press 770.
- Onagbola EO, Meyer WL, Boina DR, Stelinski LL (2008) Morphological characterization of the antennal sensilla of the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), with reference to their probable functions. Micron 39: 1184-1191.
- Wen-Xia D, Zhong-Ning Z (2006) Antennal sensilla of Microplitis mediator observed with scanning electron microscope. Acta Entomologica Sinica 49: 1054-1059.
- Keil TA (1996) Sensilla on the maxillary palps of Helicoverpa armigera caterpillars: In search of the CO(2)receptor. Tissue Cell 28: 703-717.
- Stange G, Stowe S (1999) Carbon-dioxide sensing structures in terrestrial arthropods. Microsc Res Tech 47: 416-427.
- Sukontason K, Methanitikorn R, Chaiwong T, Kurahashi H, Vogtsberger RC, et al. (2007) Sensilla of the antenna and palp of Hydrotaea chalcogaster (Diptera: Muscidae). Micron 38: 218-223.
- De Facci M, Wallén R, Hallberg E, Anderbrant O (2011) Flagellar sensilla of the eusocial gall-inducing thripsKladothrips intermedius and its kleptoparasite, Koptothrips dyskritus (Thysanoptera: Phlaeothripinae). Arthropod Struct Dev 40: 495-508.
- Onagbola EO, Fadamiro HY (2008) Scanning electron microscopy studies of antennal sensilla of Pteromalus cerealellae (Hymenoptera: Pteromalidae). Micron 39: 526-535.
- Giglio A, Ferrero EA, Enrico P, Sandro T, Tullia Z (2003) Ultrastructure and Comparative Morphology of Mouthpart Sensilla in Ground Beetle Larvae (Insecta, Coleoptera, Carabidae). Zool Anz 242: 277-292.
- Amornsak W, Cribb B, Gordh G (1998) External morphology of antennal sensilla of Trichogramma australicum Girault (Hymenoptera: Trichogrammatidae). Int J Insect Morphol Embryol27: 67-82.
- Roux O, van Baaren J, Gers C, Arvanitakis L, Legal L (2005) Antennal structure and ovipostion behavior of the Plutella xylostella specialist parasitoid: Cotesia plutellae. Microsc Res Tech 68: 36-44.
- Keil TA, Steinbrecht RA (1984) Mechanosensitive and olfactory sensilla of insects. In: King RC, Akai H (Eds.), Insect Ultrastructure, Plenum Press, NY/London 2: 477-516.
- Schafer R (1971) Antennal sense organs of the cockroach, Leucophaea maderae. J Morphol 134: 91-103.
- Loftus R (1968) The response of the antennal cold receptor of Periplaneta americanato rapid temperature changes and to steady temperature. J Comp Physiol A 59: 413-455.
- Altner H, Loftus R (1985) Ultrastructure and function of insect thermo- and hygroreceptors. Annu Rev Entomol 30: 273-295.
- Baker GT (1987) Apical sensilla on the adult and larval labial and maxillary palpi of Odontotaenius disjunctus (Illiger) (Coleoptera: Passalidae). Proceedings of the Entomological Society of Washington 89: 682-686.
- Kaissling KE (1986) Chemo-electrical transduction in insect olfactory receptors. Ann Rev Neurosci 9: 121-145.
- Isidoro N, Bartlet E, Ziesmann J, Williams IH (1998) Antennal contact chemosensilla in Psylliodes chrysocephalaresponding to cruciferous allelochemicals. Physiol Entomol 23: 131-138.
- Lopes O, Barata EN, Mustaparta H, Araújo J (2002) Fine structure of antennan sensilla basiconica and their detection of plant volatiles in the eucalyptus woodborer, Phoracantha semipunctata Fabricius(Coleopetera: Cerambycidae). Arthropod Struct Dev 31: 1-13.
- Zacharuk RY (1985) Antennal sensilla. In: Kerkut, G.A., Gilbert, L.I. (Eds.), Comparative Insect Physiology, Biochemistry and Pharmacology.
- Hu JH, Wang ZY, Sun F (2011) Anatomical organization of antennal-lobe glomeruli in males and females of the scarab beetle Holotrichia diomphalia (Coleoptera: Melolonthidae). Arthropod Structure & Development 40: 420-428.
- McKenna MC, Kear DJ, Ellsworth RA (1995) Children?s attitudes toward reading: A national survey. Reading Research Quarterly 30: 934-955.
- Pikielny CW, Hasan G, Rouyer F, Rosbash M (1994) Members of a family of Drosophila putative odorant-binding proteins are expressed in different subsets of olfactory hairs. Neuron 12: 35-49.
- Wang Q, Hasan G, Pikielny CW (1999) Preferential expression of biotransformation enzymes in the olfactory organs of Drosophila melanogaster, the antennae. J Biol Chem 274: 10309-10315.
- Moosebag (2012) "What is an axon terminal?" Answers Corporation.
- Sauber, Colleen (2008) Focus October 20-Neurobiology Visualizing the synaptic connection.
- Giovanni GC, Sachse S (2010) The Neurobiology of Olfaction (Chapter 2: Odor Coding in Insects). Menini A, editor. Boca Raton (FL): CRC Press.
- Leon M, Johnson BA (2003) Olfactory coding in the mammalian olfactory bulb. Brain Res Brain Res Rev 42: 23-32.
- Vogt RG, Prestwich GD, Lerner MR (1991) Odorant-Binding-Protein subfamilies associate with distinct classes of olfactory receptor neurons in i nsects. J Neurobiol 22: 74-84.
- Vosshall LB, Wong AM, Axel R (2000) An olfactory sensory map in the fly brain. Cell 102: 147-159.
- Hallem EA, Dahanukar A, Carlson JR (2006) Insect odor and taste receptors. Annu Rev Entomol 51: 113-135.
- Johnson BA, Leon M (2000) Modular representations of odorants in the glomerular layer of the rat olfactory bulb and the effects of stimulus concentration. J Comp Neurol 422: 496-509.
- Mombaerts P, Wang F, Dulac C, Vassar R, Chao SK, et al. (1996) The molecular biology of olfactory perception. Cold Spring Harb Symp Quant Biol 61: 135-145.
- Vosshall LB (2000) An olfactory sensory map in the fly brain. Cell 96: 725-736.
- Couto A (2005) Molecular, anatomical and functional organizations of the Drosophila olfactory system. Curr Biol 15: 1535-1547.
- Shang Y, Claridge-Chang A, Sjulson L, Pypaert M, Miesenböck G (2007) Excitatory local circuits and their implications for olfactory processing in the fly antennal lobe. Cell 128: 601-612.
- Berg BG, Almaas TJ, Bjaalie JG, Mustaparta H (1998) The macroglomerular complex of the antennal lobe in the tobacco budworm moth Heliothis virescens: specified subdivision in four compartments according to information about biologically significant compounds. J Comparat Physiol A 183: 669-682.
- Hansson BS, Ljungberg H, Hallberg E, Löfstedt C (1992) Functional specialization of olfactory glomeruli in a moth. Science 256: 1313-1315.
- Galizia C, Menzel R (2001) The role of glomeruli in the neural representation of odours: Results from optical recording studies. J Insect Physiol 47: 115-130.
- Fiala A, Spall T, Diegelmann S, Eisermann B, Sachse S, et al. (2002) Genetically expressed cameleon in Drosophila melanogaster is used to visualize olfactory information in projection neurons. Curr Biol 12: 1877-1884.
- Wang JW, Wong AM, Flores J, Vosshall LB, Axel R (2003) Two-photon calcium imaging reveals an odor-evoked map of activity in the fly brain. Cell 112: 271-282.
- Carlsson MA, Galizia CG, Hansson BS (2002) Spatial representation of odours in the antennal lobe of the moth Spodoptera littoralis (Lepidoptera: Noctuidae). Chem Senses. 27: 231-244.
- Silbering AF, Galizia CG (2007) Processing of odor mixtures in the Drosophila antennal lobe reveals both global inhibition and glomerulus-specific interactions. J Neurosci 27: 11966-11977.
- Pareto A (1972) Die zentrale Verteilung der Fühlerrafferenz bei Arbeiterinnen der Honigbiene, Apis mellifera L. Spatial distribution of sensory antennal fibres in the central nervous system of worker bees. Z Zellforsch Mikrosk Anat 131: 109-140.
- Potter SM, Zheng C, Koos DS, Feinstein P, Fraser SE, et al. (2001) Structure and emergence of specific olfactory glomeruli in the mouse. J Neurosci 21: 9713-9723.
- Rospars JP (1988) Structure and development of the insect antennodeuto-cerebral system. Int J Ins Morphol Embryol 17: 243-294.
- Hildebrand JG, Shepherd GM (1997) Mechanisms of olfactory discrimination: converging evidence for common principles across phyla. Annu Rev Neurosci 20: 595-631.
- Ignell R, Anton S, Hansson BS (2001) The antennal lobe of orthoptera ? anatomy and evolution. Brain Behav Evol 57: 1-17.
- Galizia CG, Menzel R, Holldobler B (1999) Optical imaging of odor-evoked glomerular activity patterns in the antennal lobes of the ant Camponotus rufipes. Naturwissenschaften 86: 533-537.
- Schachtner J, Schmidt M, Homberg U (2005) Organization and evolutionary trends of primary olfactory brain centers in Tetraconata (Crustacea+Hexapoda). Arthropod Struct Develop 34: 257-299.
- Karlson P, Butenandt A (1959) Pheromones (Ectohormones) in Insects. Ann Rev Entomol 4: 39-58.
- Brossut R (1997) Phéromones: La communication chimique chez les animaux. Belin : Croisée des Sciences.
- Prestwich GD, Blomquist GJ (1987) Pheromone Biochemistry. Academic Press, New York.
- Stowe MK, Tumlinson JH, Heath RR (1987) Chemical mimicry: bolas spiders emit components of moth prey species sex pheromones. Science 236: 964-967.
- Kaissling KE (1996) Peripheral mechanism of pheromone reception in moths. Chem Senses 21: 257-268.
- Vogt RG (1987) The molecular basis of pheromone reception: its influence on behavior. In: Prestwich GD and Blomquist GJ, eds. Pheromone Biochemistry. New York: Academic Press 385-431.
- Sánchez-Gracia A, Vieira FG, Rozas J (2009) Molecular evolution of the major chemosensory gene families in insects. Heredity(Edinb) 103: 208-216.
- Vogt RG (2003) Biochemical diversity of odor detection: OBPs, ODEs and SNMPs.InInsect pheromone biochemistry and molecular biology. Edited by Blomquist G, Vogt RG. San Diego: Academic Press: 391-445.
- Breer H, Krieger J, Raming K (1990) A novel class of binding proteins in the antennae of the silk moth Antheraea pernyi. Insect Biochem 20: 735-740.
- Pelosi P, Zhou JJ, Ban LP, Calvello M (2006) Soluble proteins in insect chemical communication. Cell Mol Life Sci 63: 1658-1676.
- Vieira FG, Rozas J (2011) Comparative genomics of the odorant-binding and chemosensory protein gene families across the Arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol Evol 3: 476-490.
- Law JH, Regnier FE (1971) Pheromones. Annu Rev Biochem 40: 533-548.
- Cork A (2004) Pheromone Manual. Natural Resources Institute, Chatham Maritime ME4 4TB, UK.
- Stamp N (2003) Out of the quagmire of plant defense hypotheses. Q Rev Biol 78: 23-55.
- Brown WL, Eisner T, Whittaker WH (1970) Allomones and kairomones: Transspecific chemical messengers. BioScience 20: 21-22.
- Mechaber W (2001) Interactions, Plant-Insect. Plant science.
- Tumlinson JH, Lewis WJ, Vet LEM (1993) How parasitic wasps find their hosts. Sci Am 268: 100-106.
- Marco EM, García-Gutiérrez MS, Bermúdez-Silva FJ, Moreira FA, Guimarães F, et al. (2011) Endocannabinoid system and psychiatry: in search of a neurobiological basis for detrimental and potential therapeutic effects. Front Behav Neurosci 5: 63.
- Raguso RA (2008) Wake up and smell the roses: the ecology and evolution of ?oral scent. Annu Rev Ecol Evol Syst 39: 549-569.
- Bruce TJ, Wadhams LJ, Woodcock CM (2005) Insect host location: a volatile situation. Trends Plant Sci 10: 269-274.
- Martin JP, Beyerlein A, Dacks AM, Reisenman CE, Riffell JA, et al. (2011) The neurobiology of insect olfaction: Sensory processing in a comparative context. Prog Neurobiol 95: 427-447.
- Murlis J, Elkinton JS, Carde´ RT (1992) Odor plumes and how insects use them. Annu Rev Entomol 37: 505-532.
- Frey M, Chomet P, Glawischnig E, Stettner C, Grun S, et al. (1997) Analysis of a chemical plant defense mechanism in grasses. Science 277: 696-699.
- Lichtenthaler HK, Rohmer M, Schwender J (1997) Two independent biochemical pathways for isopentenyl diphosphate and isoprenoid biosynthesis in higher plants. Physiol Plant 101: 643-652.
- Donath J, Boland W (1994) Biosynthesis of acyclic homoterpenes in higher plants parallels steroid hormone metabolism. J Plant Physiol 143: 473-478.
- Blee E (1998) Phytooxylipins and plant defense reactions. Prog Lipid Res 37: 33-72.
- Paré PW, Tumlinson JH (1999) Plant Volatiles as a Defense against Insect Herbivores. Plant Physiol 121: 325-332.
- Larsson MC, Leal WS, Hansson BS (2001) Olfactory receptor neurons detecting plant odours and male volatiles in Anomala cuprea beetles (Coleoptera : Scarabaeidae). J Insect Physiol 47: 1065-1076.
- Wall C, Perry JN (1987) Range of action of moth sex-attractant sources. Entomol Exp Appl 44: 5-14.
- Dudareva N, Pichersky E (2000) Biochemical and molecular genetic aspects of ?oral scents. Plant Physiol 122: 627-633.
- Cunningham GB, Van Buskirk R, Hodges MJ, Nevitt GA (2006) Behavioural responses of blue petrel chicks Halobaena caerulea to a food-related and novel odor in a simple wind tunnel. Antarct Sci 18: 345-352.
- Tasin M, Bäckman AC, Bengtsson M, Ioriatti C, Witzgall P (2006) Essential host plant cues in the grapevine moth. Naturwissenschaften 93: 141-144.
- Birkett MA, Bruce TJ, Martin JL, Smart LE, Oakley J, et al. (2004) Responses of female orange wheat blossom midge,Sitodiplosis mosellana, to wheat panicle volatiles. J Chem Ecol 30: 1319-1328.
- Tasin M, Bäckman AC, Coracini M, Casado D, Ioriatti C, et al. (2007) Synergism and redundancy in a plant volatile blend attracting grapevine moth females. Phytochemistry 68: 203-209.
- Kogan M (1998) Integrated pest management: historical perspectives and contemporary developments. Annu Rev Entomol 43: 243-270.
- Turlings TC, Tumlinson JH, Heath RR, Proveaux AT, Doolittle RE (1991) Isolation and identification of allelochemicals that attract the larval parasitoid, Cotesia marginiventris (Cresson), to the microhabitat of one of its hosts. J Chem Ecol 17: 2235-2251.
- Dicke M, Van Baarlen P, Wessels R, Dijkman H (1993) Herbivory induces systemic production of plant volatiles that attract predators of the herbivore: extraction of endogenous elicitor. J Chem Ecol 19: 581-599.
- Birkett MA, Chamberlain K, Guerrieri E, Pickett JA, Wadhams LJ, et al. (2003) Volatiles from whiteflyinfested plants elicit a host-locating response in the parasitoid, Encarsia formosa. J Chem Ecol 29: 1589-1600.
- Wei J, Wang L, Zhu J, Zhang S, Nandi OI, et al. (2007) Plants attract parasitic wasps to defend themselves against insect pests by releasing hexenol. PLoS One 2: e852.
- Hunter MD (2002) A breath of fresh air: beyond laboratory studies of plant volatile-natural enemy interactions. Agric Forest Entomol 4: 81-86.
- James DG (2003) Field evaluation of herbivore-induced plant volatiles as attractants for beneficial insects: methyl salicylate and the green lacewing, Chrysopa nigricornis. J Chem Ecol 29: 1601-1609.
- James DG (2005) Further field evaluation of synthetic herbivoreinduced plant volatiles as attractants for beneficial insects. J Chem Ecol 31: 481-495.
- Schuster DJ (2007) Suppression of Anthonomus eugenii (Coleoptera: Curculionidae) pepper fruit infestation with releases of Catolaccus hunteri (Hymenoptera: Pteromalidae). Biocontrol Sci Technol 17: 345-351.
- Butenandt A, Beckmann R, Stamm D, Hecker E (1959) Über den Sexuallockstoff den Seidenspinners Bombyx mori. Reindarstellung und Konstitution. Z Naturforsch 14b: 283-284.
- Leos-Martinez J, Granovsky TA, Williams HJ, Vinson SB, Burkholder WE (1987) Pheromonal trapping methods for lesser grain borer, Rhyzopertha dominica (Coleoptera: Bostrichidae). Environ Entomol 16: 747-751.
- Weinzierl R, Henn T, Koehler PG, Tucker CL (2005) Insect attractants and traps. University of Florida, IFAS Extension.
- Phillips TW (1997) Semiochemicals of stored-product insects: research and applications. J stored Prod Res 33: 17-30.
- Cook SM, Khan ZR, Pickett JA (2007) The use of push-pull strategies in integrated pest management. Annu Rev Entomol 52: 375-400.
- Pajares JA, Ibeas F, Díez JJ, Gallego D (2004) Attractive responses byMonochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. J Applied Entomol 128: 633-638.
- Khan ZR, Midega CA, Bruce TJ, Hooper AM, Pickett JA (2010) Exploiting phytochemicals for developing a ?push?pull? crop protection strategy for cereal farmers in Africa. J Exp Bot 61: 4185-4196.
- Landolt JP (1997) Sex attractant and aggregation pheromones of male phytophagous insects. Am Entomol 43: 12-22.
- Asanov K (1980) Predators and parasites of the lesser grain borer. Zashchita Rastenii 5: 23.
- Dal Bello G, Padin S, López Lastra C, Fabrizio M (2001) Laboratory evaluation of chemical-biological control of the rice weevil (Sitophilus oryzae L.) in stored grains. J Stored Prod Res 3: 77-81.
- Begon M, Townsend C, Harper J (1996) Ecology: Individuals, populations and communities (Third edition). Blackwell Science, London.
- Brown TA (2010) Gene Cloning and DNA Analysis: An Introduction, 6th Edition. Wiley-Blackwell.
- Bruin J, Sabelis MW, Dicke M (1995) Do plants tap SOS signals from their infested neighbours? Trends Ecol Evol 10: 167-170.
- Buck L, Axel R (1991) odorant receptors and the organization of the olfactory system. Cell 65: 175-187.
- Calvello M, Brandazza A, Navarrini A, Dani FR, Turillazzi S, et al. (2005) Expression ofodorant-binding proteins and chemosensory proteins in some Hymenoptera. Insect Biochem Mol Biol 35: 297-307.
- Debanne D, Campanac E, Bialowas A, Carlier E, Alcaraz G (2011) Axon Physiology. Physiol Rev 91: 555-602.
- Fan J, Francis F, Liu Y, Chen JL, Cheng DF (2011) An overview of odorant-binding protein functions in insect peripheral olfactory reception. Genet Mol Res 10: 3056-3069.
- Cunningham JP, Moore CJ, Zalucki MP, Cribb BW (2006) Insect odour perception: recognition of odour components by flower foraging moths. Proc R Soc B 273: 2035-2040.
- Kandel ER, Schwartz JH, Jessell TM (2000) Principles of Neural Science (4th ed). New York: McGraw-Hill.
- Karban R, Maron J, Felton GW, Ervin G, Eichenseer H (2003) Herbivore damage to sagebrush induces resistance in wild tobacco: evidence for leaves dropping between plants. Oikos 100: 325-332.
- Spletter ML, Luo L (2008) A new family of Odorant Receptors in Drosophila. Cell 136: 23-25.
- Mattiacci L, Dicke M, Posthumus MA (1994) Induction of parasitoid attracting synomone in brussels sprouts plants by feeding of Pieris brassicaelarvae: role of mechanical damage and herbivore elicitor. J Chem Ecol 20: 2229-2247.
- Müller U, Hildebrandt H (2002) Nitric oxide/cGMP-mediated protein kinase A activation in the antennal lobes plays an important role in appetitive reflex habituation in the honeybee. J Neurosci 22: 8739-8747.
- Price PW, Slobodchikoff CN, Gaud WS (1984) A new ecology. Wiley-Interscience.
- Schacter DL, Gilbert DT, Wegner DM (2011) Psychology second edition. New York, NY: Worth.
- Tavosanis G (2012) Dendritic structural plasticity. Dev Neurobiol 72 : 73-86.
- Urbanska M, Blazejczyk M, Jaworski J (2008) Molecular basis of dendritic arborization. Acta Neurobiol Exp 68: 264-288.
- Vogt RG (2005) Molecular Basis of Pheromone Detection in Insects. In Comprehensive Insect Physiology, Biochemistry, Pharmacology and Molecular Biology. Gilbert LI, Iatro K, Gill S. Endocrinology 3: 753-804.
- Wakil W, Riasat T, Ashfaq M (2012) Residual efficacy of thiamethoxam, Beauveria bassiana (Balsamo) Vuillemin, and diatomaceous earth formulation against Rhyzopertha dominica F. (Coleoptera: Bostrychidae). J Pest Sci 85: 341-350.
- http://insects.about.com/od/behaviorcommunication/f/how-insects-taste.htm
- https://www.msu.edu/user/miller20/carmona.htm
- http://en.wikipedia.org/wiki/Olfactory_receptor
- http://www.cals.ncsu.edu/course/ent425/tutorial/chemo.html